
| 產品編號 | bs-0233R |
| 英文名稱 | Rabbit Anti-TERT antibody |
| 中文名稱 | 端粒酶反轉錄酶抗體 |
| 別 名 | Telomerase catalytic subunit; EST2; hEST2; TCS1; Telomerase associated protein 2; Telomere Reverse Transcriptase; TP2; TRT; Telomerase reverse transcriptase; Telomerase Catalytic Subunit; Telomerase-associated protein 2; TERT_HUMAN. |
 |
Specific References (16) | bs-0233R has been referenced in 16 publications.
|
| 研究領域 | 腫瘤 細胞生物 免疫學 神經生物學 信號轉導 細胞凋亡 細胞周期蛋白 激酶和磷酸酶 |
| 抗體來源 | Rabbit |
| 克隆類型 | Polyclonal |
| 交叉反應 | Human,Mouse,Rat |
| 產品應用 | WB=1:500-2000,IHC-P=1:100-500,IHC-F=1:100-500,Flow-Cyt=1μg/Test,ICC/IF=1:100,IF=1:100-500
not yet tested in other applications. optimal dilutions/concentrations should be determined by the end user. |
| 理論分子量 | 125kDa |
| 細胞定位 | 細胞核 細胞漿 |
| 性 狀 | Liquid |
| 濃 度 | 1mg/ml |
| 免 疫 原 | KLH conjugated synthetic peptide derived from human TERT: 601-750/1132 |
| 亞 型 | IgG |
| 純化方法 | affinity purified by Protein A |
| 緩 沖 液 | 0.01M TBS (pH7.4) with 1% BSA, 0.02% Proclin300 and 50% Glycerol. |
| 保存條件 | Shipped at 4℃. Store at -20℃ for one year. Avoid repeated freeze/thaw cycles. |
| 注意事項 | This product as supplied is intended for research use only, not for use in human, therapeutic or diagnostic applications. |
| PubMed | PubMed |
| 產品介紹 |
Telomerase is a ribonucleoprotein polymerase that maintains telomere ends by addition of the telomere repeat TTAGGG. The enzyme consists of a protein component with reverse transcriptase activity, encoded by this gene, and an RNA component which serves as a template for the telomere repeat. Telomerase expression plays a role in cellular senescence, as it is normally repressed in postnatal somatic cells resulting in progressive shortening of telomeres. Deregulation of telomerase expression in somatic cells may be involved in oncogenesis. Studies in mouse suggest that telomerase also participates in chromosomal repair, since de novo synthesis of telomere repeats may occur at double-stranded breaks. Alternatively spliced variants encoding different isoforms of telomerase reverse transcriptase have been identified; the full-length sequence of some variants has not been determined. Alternative splicing at this locus is thought to be one mechanism of regulation of telomerase activity. [provided by RefSeq, Jul 2008]. Function: Telomerase is a ribonucleoprotein enzyme essential for the replication of chromosome termini in most eukaryotes. Active in progenitor and cancer cells. Inactive, or very low activity, in normal somatic cells. Catalytic component of the teleromerase holoenzyme complex whose main activity is the elongation of telomeres by acting as a reverse transcriptase that adds simple sequence repeats to chromosome ends by copying a template sequence within the RNA component of the enzyme. Catalyzes the RNA-dependent extension of 3'-chromosomal termini with the 6-nucleotide telomeric repeat unit, 5'-TTAGGG-3'. The catalytic cycle involves primer binding, primer extension and release of product once the template boundary has been reached or nascent product translocation followed by further extension. More active on substrates containing 2 or 3 telomeric repeats. Telomerase activity is regulated by a number of factors including telomerase complex-associated proteins, chaperones and polypeptide modifiers. Modulates Wnt signaling. Plays important roles in aging and antiapoptosis. Subunit: Homodimer; dimerization is required to produce a functional complex. Oligomer; can form oligomers in the absence of the telomerase RNA template component (TERC). Catalytic subunit of the telomerase holoenzyme complex composed minimally of TERT and TERC. The telomerase complex is composed of TERT, DKC1, WDR79/TCAB1, NOP10, NHP2, GAR1, TEP1, EST1A, POT1 and a telomerase RNA template component (TERC). The molecular chaperone HSP90/P23 complex is required for correct assembly and stabilization of the active telomerase. Interacts directly with HSP90A and PTGES3. Interacts with HSPA1A; the interaction occurs in the absence of TERC and dissociates once the complex has formed. Interacts with RAN; the interaction promotes nuclear export of TERT. Interacts with XPO1. Interacts with PTPN11; the interaction retains TERT in the nucleus. Interacts with NCL (via RRM1 and C-terminal RRM4/Arg/Gly-rich domains); the interaction is important for nucleolar localization of TERT. Interacts with SMARCA4 (via the bromodomain); the interaction regulates Wnt-mediated signaling. Interacts with MCRS1 (isoform MCRS2); the interaction inhibits in vitro telomerase activity. Interacts with PIF1; the interaction has no effect on the elongation activity of TERT. Interacts with PML; the interaction recruits TERT to PML bodies and inhibits telomerase activity. Subcellular Location: Nucleus, nucleolus. Nucleus, nucleoplasm. Nucleus. Chromosome, telomere. Cytoplasm. Nucleus, PML body. Note=Shuttling between nuclear and cytoplasm depends on cell cycle, phosphorylation states, transformation and DNA damage. Diffuse localization in the nucleoplasm. Enriched in nucleoli of certain cell types. Translocated to the cytoplasm via nuclear pores in a CRM1/RAN-dependent manner involving oxidative stress-mediated phosphorylation at Tyr-707. Dephosphorylation at this site by SHP2 retains TERT in the nucleus. Translocated to the nucleus by phosphorylation by AKT. Tissue Specificity: Expressed at a high level in thymocyte subpopulations, at an intermediate level in tonsil T-lymphocytes, and at a low to undetectable level in peripheral blood T-lymphocytes. Post-translational modifications: Ubiquitinated, leading to proteasomal degradation. Phosphorylation at Tyr-707 under oxidative stress leads to translocation of TERT to the cytoplasm and reduces its antiapoptotic activity. Dephosphorylated by SHP2/PTPN11 leading to nuclear retention. Phosphorylation by the AKT pathway promotes nuclear location. DISEASE: Note=Activation of telomerase has been implicated in cell immortalization and cancer cell pathogenesis. Defects in TERT are associated with susceptibilty to aplastic anemia (AA) [MIM:609135]. AA is a rare disease in which the reduction of the circulating blood cells results from damage to the stem cell pool in bone marrow. In most patients, the stem cell lesion is caused by an autoimmune attack. T-lymphocytes, activated by an endogenous or exogenous, and most often unknown antigenic stimulus, secrete cytokines, including IFN-gamma, which would in turn be able to suppress hematopoiesis. Note=Genetic variations in TERT are associated with coronary artery disease (CAD). Defects in TERT are the cause of dyskeratosis congenital autosomal dominant type 2 (DKCA2) [MIM:613989]. A rare multisystem disorder caused by defective telomere maintenance. It is characterized by progressive bone marrow failure, and the clinical triad of reticulated skin hyperpigmentation, nail dystrophy, and mucosal leukoplakia. Common but variable features include premature graying, aplastic anemia, low platelets, osteoporosis, pulmonary fibrosis, and liver fibrosis among others. Early mortality is often associated with bone marrow failure, infections, fatal pulmonary complications, or malignancy. Defects in TERT are the cause of dyskeratosis congenital autosomal recessive type 4 (DKCB4) [MIM:613989]. A rare multisystem disorder caused by defective telomere maintenance. It is characterized by progressive bone marrow failure, and the clinical triad of reticulated skin hyperpigmentation, nail dystrophy, and mucosal leukoplakia. Common but variable features include premature graying, aplastic anemia, low platelets, osteoporosis, pulmonary fibrosis, and liver fibrosis among others. Early mortality is often associated with bone marrow failure, infections, fatal pulmonary complications, or malignancy. Defects in TERT are a cause of susceptibility to pulmonary fibrosis idiopathic (IPF) [MIM:178500]. Pulmonary fibrosis is a lung disease characterized by shortness of breath, radiographically evident diffuse pulmonary infiltrates, and varying degrees of inflammation and fibrosis on biopsy. It results in acute lung injury with subsequent scarring and endstage lung disease. Similarity: Belongs to the reverse transcriptase family. Telomerase subfamily. Contains 1 reverse transcriptase domain. SWISS: O14746 Gene ID: 7015 Database links: Entrez Gene: 7015 Human Entrez Gene: 21752 Mouse SwissProt: O14746 Human SwissProt: O70372 Mouse
端粒酶是一種依賴RNA的DNA聚合酶,催化合成端粒體的DNA重復序列,并引導端粒添加到染色體的尾端,對維持染色體的長度、調節細胞增殖和凋亡起重要作用。 端粒酶主要存在于惡性腫瘤之中,一般在大多數正常組織中沒有活性或活性極低,同時由于端粒酶在惡性腫瘤的發展中起著關鍵作用,所以通過各種途徑抑制端粒酶的活性可能有效地抑制大多數腫瘤的生長,而對大多數正常細胞沒有影響。這種抑制作用可以通過直接抑制端粒酶活性、抑制端粒酶RNA或端粒酶蛋白成分以及誘導腫瘤細胞發生分化等方法實現。 端粒酶與惡性腫瘤之間令人驚異的相關性使它在腫瘤的診斷和治療上有望成為行之有效的新的靶目標。 |
| 產品圖片 |
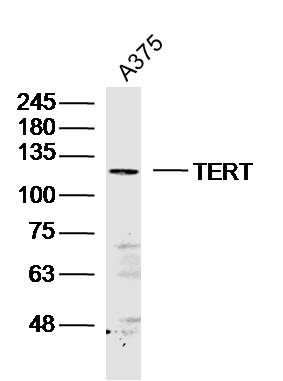
Sample: A375 (human)cell Lysate at 40 ug
Primary: Anti-TERT (bs-0233R)at 1/300 dilution
Secondary: IRDye800CW Goat Anti-Rabbit IgG at 1/20000 dilution
Predicted band size: 125kD
Observed band size: 125 kD
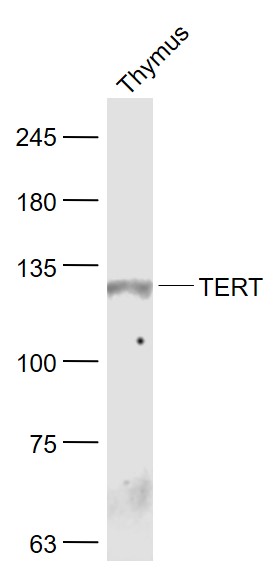
Sample:
Thymus (Mouse) Lysate at 40 ug
Primary: Anti- TERT (bs-0233R) at 1/1000 dilution
Secondary: IRDye800CW Goat Anti-Rabbit IgG at 1/20000 dilution
Predicted band size: 125 kD
Observed band size: 125 kD
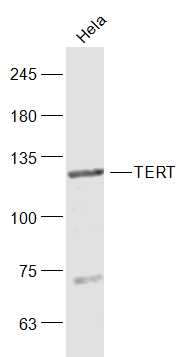
Sample:
Hela(Human) Cell Lysate at 30 ug
Primary: Anti-TERT (bs-0233R) at 1/1000 dilution
Secondary: IRDye800CW Goat Anti-Rabbit IgG at 1/20000 dilution
Predicted band size: 125 kD
Observed band size: 125 kD

Tissue/cell: human stomach carcinoma; 4% Paraformaldehyde-fixed and paraffin-embedded;
Antigen retrieval: citrate buffer ( 0.01M, pH 6.0 ), Boiling bathing for 15min; Block endogenous peroxidase by 3% Hydrogen peroxide for 30min; Blocking buffer (normal goat serum,C-0005) at 37℃ for 20 min;
Incubation: Anti-TERT Polyclonal Antibody, Unconjugated(bs-0233R) 1:400, overnight at 4°C, followed by conjugation to the secondary antibody(SP-0023) and DAB(C-0010) staining
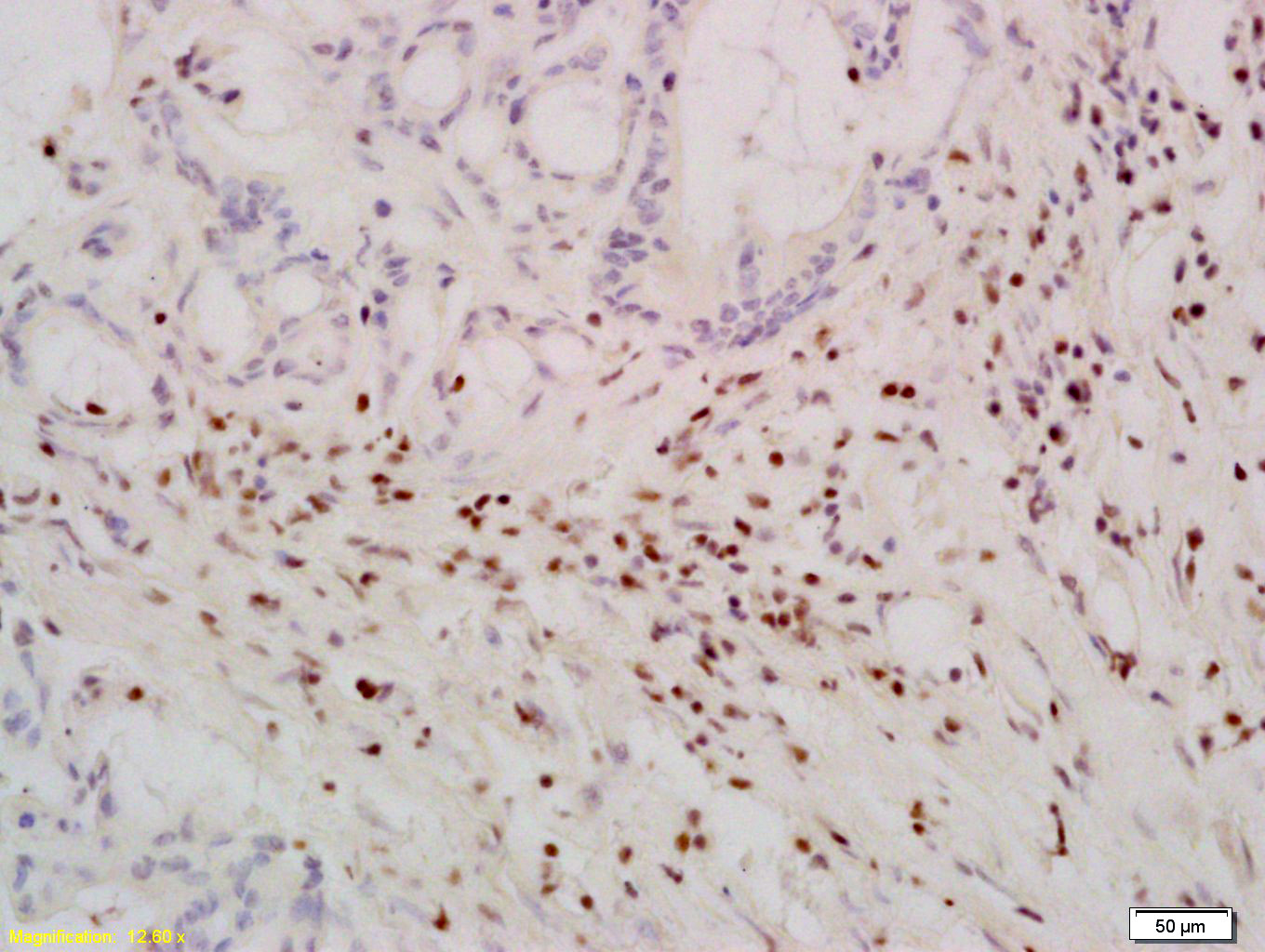
Tissue/cell: human colon carcinoma; 4% Paraformaldehyde-fixed and paraffin-embedded;
Antigen retrieval: citrate buffer ( 0.01M, pH 6.0 ), Boiling bathing for 15min; Block endogenous peroxidase by 3% Hydrogen peroxide for 30min; Blocking buffer (normal goat serum,C-0005) at 37℃ for 20 min;
Incubation: Anti-TERT Polyclonal Antibody, Unconjugated(bs-0233R) 1:200, overnight at 4°C, followed by conjugation to the secondary antibody(SP-0023) and DAB(C-0010) staining
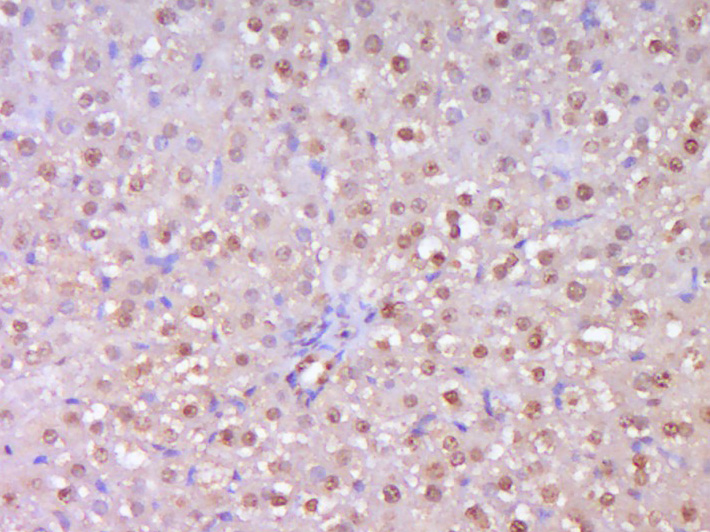
Paraformaldehyde-fixed, paraffin embedded (Rat liver); Antigen retrieval by boiling in sodium citrate buffer (pH6.0) for 15min; Block endogenous peroxidase by 3% hydrogen peroxide for 20 minutes; Blocking buffer (normal goat serum) at 37°C for 30min; Antibody incubation with (TERT) Polyclonal Antibody, Unconjugated (bs-0233R) at 1:400 overnight at 4°C, followed by operating according to SP Kit(Rabbit) (sp-0023) instructions and DAB staining.
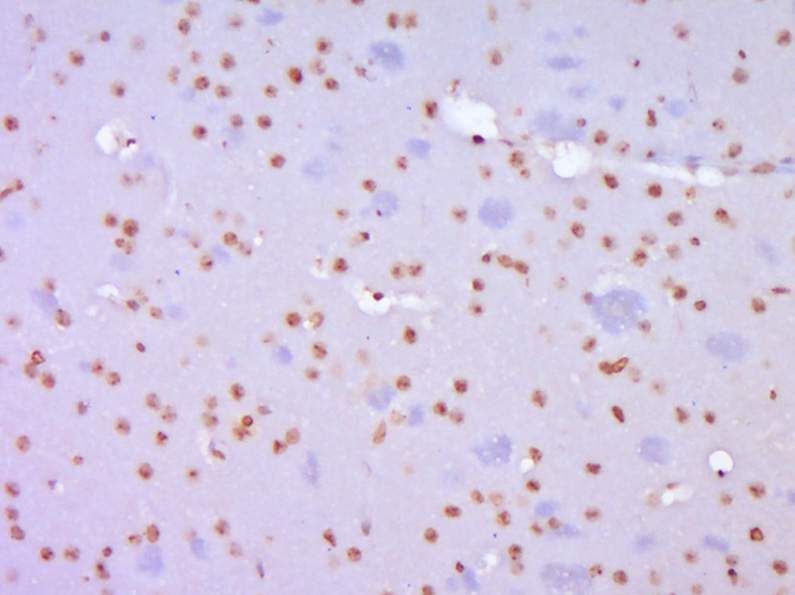
Paraformaldehyde-fixed, paraffin embedded (Mouse brain); Antigen retrieval by boiling in sodium citrate buffer (pH6.0) for 15min; Block endogenous peroxidase by 3% hydrogen peroxide for 20 minutes; Blocking buffer (normal goat serum) at 37°C for 30min; Antibody incubation with (TERT) Polyclonal Antibody, Unconjugated (bs-0233R) at 1:400 overnight at 4°C, followed by operating according to SP Kit(Rabbit) (sp-0023) instructions and DAB staining.
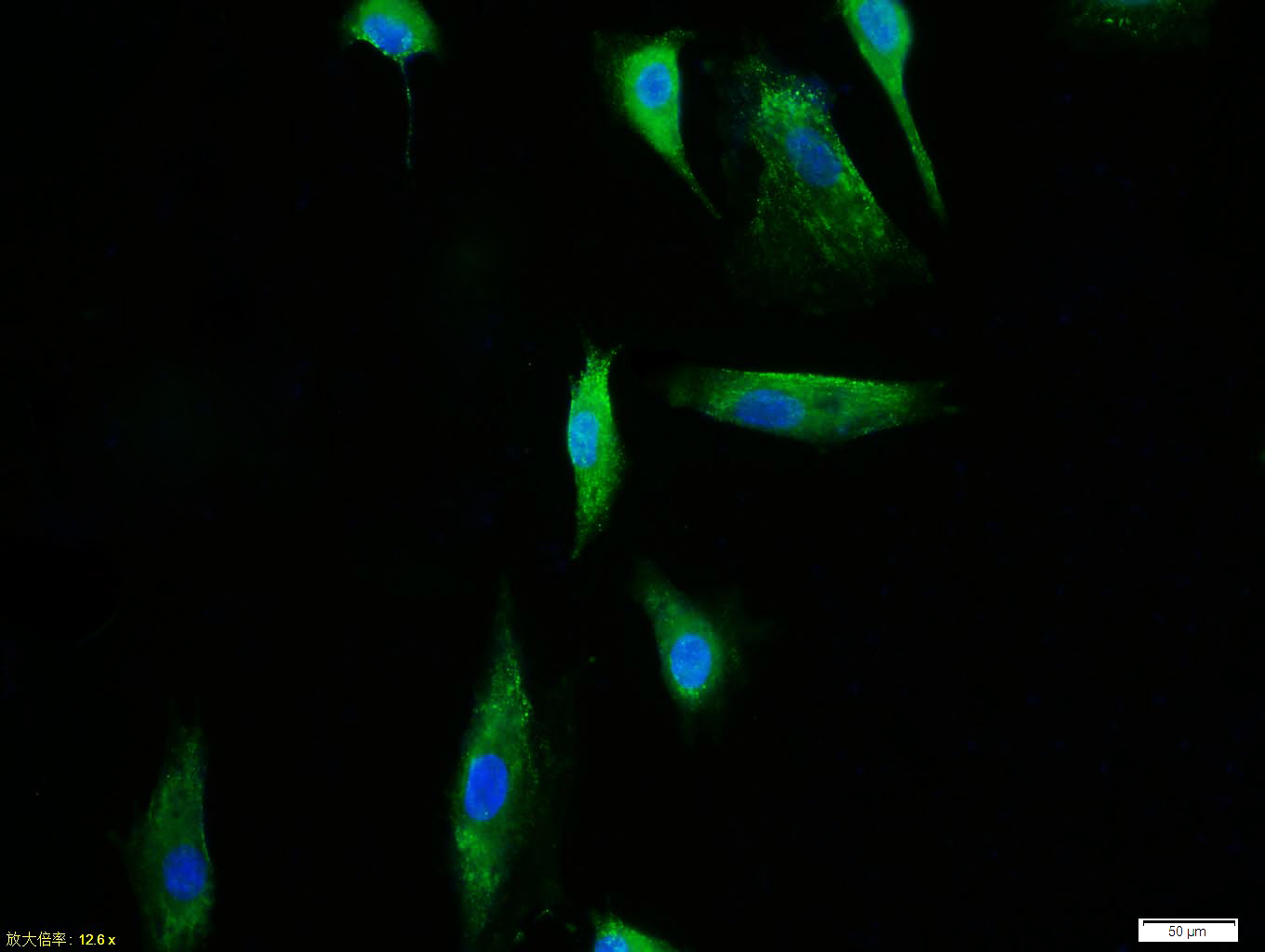
Tissue/cell:A549 cell; 4% Paraformaldehyde-fixed; Triton X-100 at room temperature for 20 min; Blocking buffer (normal goat serum,C-0005) at 37°C for 20 min; Antibody incubation with (TERT) polyclonal Antibody, Unconjugated (bs-0233R) 1:100, 90 minutes at 37°C; followed by a FITC conjugated Goat Anti-Rabbit IgG antibody at 37°C for 90 minutes, DAPI (blue, C02-04002) was used to stain the cell nuclei.
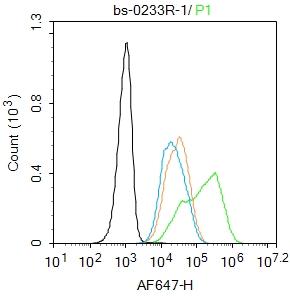
Blank control:Hela.
Primary Antibody (green line): Rabbit Anti-TERT antibody (bs-0233R)
Dilution: 1μg /10^6 cells;
Isotype Control Antibody (orange line): Rabbit IgG .
Secondary Antibody : Goat anti-rabbit IgG-AF647
Dilution: 1μg /test.
Protocol
The cells were fixed with 4% PFA (10min at room temperature)and then permeabilized with 90% ice-cold methanol for 20 min at-20℃.The cells were then incubated in 5%BSA to block non-specific protein-protein interactions for 30 min at room temperature .Cells stained with Primary Antibody for 30 min at room temperature. The secondary antibody used for 40 min at room temperature. Acquisition of 20,000 events was performed.
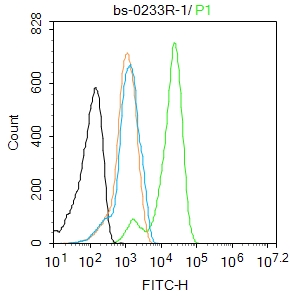
Blank control: K562.
Primary Antibody (green line): Rabbit Anti-TERT antibody (bs-0233R)
Dilution: 1μg /10^6 cells;
Isotype Control Antibody (orange line): Rabbit IgG .
Secondary Antibody : Goat anti-rabbit IgG-FITC
Dilution: 1μg /test.
Protocol
The cells were fixed with 4% PFA (10min at room temperature)and then permeabilized with 90% ice-cold methanol for 20 min at -20℃.The cells were then incubated in 5%BSA to block non-specific protein-protein interactions for 30 min at room temperature .Cells stained with Primary Antibody for 30 min at room temperature. The secondary antibody used for 40 min at room temperature. Acquisition of 20,000 events was performed.
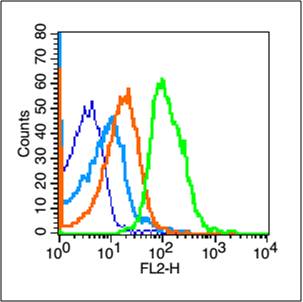
Blank control (blue line): Mouse thymus cells (blue).
Primary Antibody (green line): Rabbit Anti-TERT antibody (bs-0233R)
Dilution: 1μg /10^6 cells;
Isotype Control Antibody (orange line): Rabbit IgG .
Secondary Antibody (white blue line): Goat anti-rabbit IgG-PE
Dilution: 1μg /test.
Protocol
The cells were fixed with 70% methanol (Overnight at 4℃) and then permeabilized with 90% ice-cold methanol for 20 min at -20℃. Cells stained with Primary Antibody for 30 min at room temperature. The cells were then incubated in 1 X PBS/2%BSA/10% goat serum to block non-specific protein-protein interactions followed by the antibody for 15 min at room temperature. The secondary antibody used for 40 min at room temperature. Acquisition of 20,000 events was performed.
|
| 1、抗體溶解方法 | |
| 2、抗體修復方式 | |
| 3、常用試劑的配制 | |
| 4、免疫組化操作步驟 | |
| 5、免疫組化問題解答 | |
| 6、Western Blotting 操作步驟 | |
| 7、Western Blotting 問題解答 | |
| 8、關于肽鏈的設計 | |
| 9、多肽的溶解與保存 | |
| 10、酶標抗體效價測定程序 | |






